Способы противодействия на эпигеномном уровне прогрессированию раковых и сенесцентных клеток

Способы противодействия раковым и сенесцентным клеткам с помощью подавления провоспаления и гликолиза
Уровень мелатонина снижает гормон стресса - кортизол, а также провоспалительных интерлейкинов, которые всегда сопровождают и стимулируют онкологию. Кортизол, провоспалительные интерлейкины, раковая кислотная интоксикация – все они элементы клеточного и иммунного стресса - снижают резко уровень выработки мелатонина и стимулируют рак. И наоборот, отсутствие мелатонина – усиливает их действие.
Мелатонин можно принимать параллельно с нашей Программой по 3-5 мг. Большие дозы антиоксидантов и противовоспалительных не нужны.
При этом одностороннее повышение мелатонина ничего не даёт. Уберём хронический провоспалительный клеточный стресс – это уже первый шаг к торможению развития онкологии. Задача одновременно снижать! уровень кортизола и интерлейкинов, но повышать! мелатонина. Часто для этого надо разобраться в причинах почему происходит такой перекос, искать корни откуда зарождается перекос… и их устранять. Простому человеку не дано разобраться в этой массе тонкостей. Поэтому задача хотя бы гасить клеточный эпигенетический стресс и всё множество его компонентов.
Следует понимать, что регулировка раковых опухолей происходит по принципу маятника: одни и те же препараты, например антиоксиданты, в одних случаях действуют положительно и тормозят опухоль, а в других – отрицательно. Всё зависит от того какой рычаг маятника перевесит. В большинстве случаев при прогрессирующем раке все односторонние методики по его гашению подобны подбросу дров в огонь. Важно устранять более радикальные разжигающие первопричины – клеточный стресс! Тогда те же средства, что разжигали начнут подавлять опухоль.
К противовоспалительным относятся: Куркумин-Адванс, Витамин D3 Экспресс Форте, Фульвовый комплекс Сrystal, Фуллерен C60…
К антигликолизным относятся: Магний Экспресс Форте, NMN (Никотинамидмононуклеотид), Янтарная кислота… - закажите у нас; рассол помидорный или огуречный.
Рак - это не мутация, а перекос в эпигеномных регулировках-настройках. При этом следует учитывать, что перекос может зависеть как от гиперкомпактизации хроматина (патологическая «невидимость генов»), так и разрыхления хроматина (вскрытие той части хромосомы, которая не должно быть «видимой»).
В норме на пути всем этим перекосам стоит страховочный белок, который может служить мощным препятствием для упаковки ДНК и формировать “жесткие границы”. Он работает по принципу «колок»*-настройщиков. Известно, что РНК-полимеразы, продвигаясь по активному гену, «сметают» большинство белков на своем пути, но не могут разорвать особые структуры — когезиновые кольца. Но это правило работает в нормальных условиях регулировок, когда они не выходят за пределы коридора оптимума гомеостазов. Тем не менее, очевидно при онкологии создаются именно такие условия выхода за пределы коридора гармонии биохимических процессов, когда вся системы выходит далеко за эти пределы, что создаёт условия для «прорыва». При этом РНК-полимеразы «сметают» не просто большинство белков на своём пути, но и разрывают особые структуры — когезиновые кольца. Разрыв-прорыв когезиновых «ограничителей-тормозов» ведёт к изменению функции колок, когда пропадает «настройка-натяжка струн», а система «вскрывается» над зоной курирования хроматина, ответственной за регулировку репликаций. Конечно, такое становится возможным в особых провоспалительных условиях среды для клеток.
Регулировка дисклаймерного* статуса клеток с помощью метода полуавтономии. Именно эту среду и надо радикально поменять, что не возможно без полуавтономного длительного питания (точнее фактически больше голодание чем питание), чтобы заставить глубинно перестроить энергетику клеток. Чистый голод не допустим! так как усиливает стресс и кортизол. Но не полноценная пища тоже будет провоцировать опухоль и переводить её на гликолиз, то есть это другая крайность. Только биологически высоко полноценная пища, но обязательно на фоне критического! недостатка калорийности.
Препараты и методы для снижения провоспаления.
Есть травы и препараты, которые снижают кортизол, например GABA/ГАБА — Экспресс-Форте.
Безопасный природный транквилизатор – есть у нас; а также помогает 1-2-х разовое питание – снижает кортизол.
Что такое сбросить КЭШ и какую роль в этом играет Полуавтономное питание?
Суть полуавтономного питания. Оно подразумевает обязательное сочетание ряда методик, которые проводят на фоне резкого снижения калорий – вплоть до 400-600 ккал в день, но полноценными продуктами: яйца, рыба, икра, молоки рыб, гречка, горошек, спирулина, костный мозг... Важно чтобы эта пища состояла из живых продуктов для активирования эффекта парабиогенеза*. Смотрите ниже рекомендации по 1-разовому питанию! Только такой принцип питания запустит механизмы полуавтономного метаболизма. Точнее перезапустит настройки сдвинутого эпигенома в норму.
Полуавтономное питание по сути заменяет Фазу 1 нашей методики. Его следует проводить около 3 месяцев подряд, чтобы возможно было отследить динамику лечения. Затем надо будет прислать отчёт. При этом важно проводить самоконтроль за своим самочувствием: улучшение всех этих параметров указывает на то, что идёт положительная динамика.
Критерии положительной динамики:
- улучшение самочувствия
- повышение работоспособности
- ослабление онкологических болей
- снижение температуры
- уменьшение аутоинтоксикаций и провоспаления
- размягчение или уменьшение опухоли
- отёки
Первый месяц пока организм адаптируется к нему возможно похудение, но в дальнейшем вес почти сам восстанавливается из-за перехода на экономный режим питания и самопожирания опухоли. Процесс самопоедания опухоли возможен только в условиях полного отсутствия стресса!, но на обязательном фоне безстрессового «полуголодания», так как любой голод – это стресс и провоспаления, которые стимулируют опухоль! Здесь нужна гибкая система компромиссов: между антистрессом и голоданием. Такое возможно только в условиях балансирования, маневрирования: голод, но не голод, точнее автономный режим, когда организм длительно способен существовать преимущественно за счёт внутренних ресурсов. Только так можно медленно заморить раковую опухоль, но не повредить организму, а главное не только не спровоцировать стрессы, а наоборот от них (онкологических стрессов самопровоцирования) избавиться.
Реализуют, направляют этот процесс только иммунные клетки (макрофаги и лимфоциты). И они должны быть здоровыми, полноценными, для них мы поддерживаем на минимуме полноценную пищу! Любое как перерождение (онкологическое или дегенеративное), так и восстановление происходит только при участии иммунных клеток. Задача возродить изначальную Жизненную Силу макрофагов, «перепрограммировать» их. В этом случае их целью становится не столько подавить опухоль за счёт макрофагии клетками киллерами или стимулирования апоптоза в онкоклетках, а перенаправить, перезапустить эти клетки на путь ремодификации в норму, снять эпигенетическую программу «зависания». Следует понимать, что в условиях стресса (скрытое провоспаление) и калорийной насыщенности организма иммунные клетки наоборот помогают раковым клеткам. Палка о двух концах… Задача возрождать полноценность и силу этих иммунных клеток-резидентов, киллеров = убийц для рака. Пробудить их воинствующую силу надо путём предварительного устранения клеточного стресса. В итоге такой комплексный подход изменит онкологический эпигенетический статус онкоклеток и рано или поздно выведет их из цейтнота делеций (разрывы ДНК по причине эпигенетических разрегулировок настроек) на путь самоуничтожения (апоптоз) или их уничтожат макрофаги: иммунные клетки-киллеры (NK-клетки), или в них будет происходить реверсия, репрограммирование, ремодификация в нормальные. Очевидно онкология с делециями* (ошибочно их принимают за мутации) будет на много сложнее переводиться в норму.
Роль полуавтономии как механизма сбрасывания КЭШа. Дословно очистить кэш означает удаление временных данных, которые хранятся в кэше (временном хранилище) программы, браузера, операционной системы или приложения. Кэш позволяет ускорить доступ к часто используемым файлам, так как система берёт их из локальной памяти, а не загружает с сервера. Это снижает нагрузку на основное хранилище, ускоряет загрузку данных и повышает производительность системы. Но этот же кэш засоряет и перегружает систему. По сути, эпигеном это система локальной памяти типа скриптов, ссылок…, а точнее паттернов, которая по аналогии с компьютером приобретается в ходе его работы, а в живых системах - в ходе существования клеток в виде паттернов на эпигеноме. Эпигеном можно сравнивать с браузером, который целенаправленно (как библиотекарь) обеспечивает доступ к нужным интернет данным, а в клетке – к нужным данным генома. Автор под избыточным кэшем понимает «зависание системы» из-за перегрузки в эпигенетических настройках, а также механизмы-настройки ведущие к сенесценту клеток или онкологизации. Без снятия этого «груза», а иногда и информационного «мусора» невозможно запустить механизмы реювентализации. Таким образом, понимается необходимость снятия «зависания» в дисклаймере* (за пределами гомеостаза, то есть зоны где не достают сигналы регулировок), а также инициация, перезапуск более высоких потентностей и вплоть до реювентализации. Рычагов воздействия должно быть два: воздействие на дисклаймер (эпигеном), а также на клеточные органеллы и среду (мантию). В первом случае эту задачу можно решить с помощью метода полуавтономии, а в другом – с помощью аутофагии. Следовательно это два рычага воздействия на одну систему с единой целью её вывести (из зоны энтроукта = хаоса и дисклаймера) в зону гомеостаза.
До сих научный мир даёт объяснения сенесценту, утверждая, что якобы за долгие годы в ДНК накапливаются ошибки и поломки, которые приводят к износу внутренних органов. Но автор считает это ошибочной научной доктриной и даёт этому свою трактовку: это всего лишь направленный «сбой», а точнее смещение эпигенетических настроек или заранее заданное «окно возможностей», детерминация, причем точнее даже не «сбой», а коммитированно (предписано, предрасположено) направленный процесс. Ну почему у короткоживущих животных (мыши, полёвки) за 2 года может накопиться такое же критическое количество «поломок», как и у долгоживущих землекопов, которые живут 40 лет. Есть долгожители как черепахи, акулы доживающие до 200 – 500 лет. У растений вообще нет механизма сенесцента и некоторые виды могут существовать тысячелетия (возраст рощи тополя осинолистного 80 тысяч лет). У термитов рабочие особи живут месяцы, а матки – десятилетия! Разница в сто раз. В пределах одного вида такая огромная разница в продолжительности жизни у различных его фенотипов! Природа фенотипов связана не с морфогенезом, а с эпигенезом, Так что теория накопления поломок и ошибок не состоятельна! Эпигенез – это по сути КЭШ, осуществляемый через МалыйБиоКристалл. Следовательно КЭШом следует считать те механизмы эпигеномных настроек, которые могут укорачивать или продлевать продолжительность жизни.
Возникает вопрос: разница в продолжительности жизни у мыши и землекопа только в различных настройках эпигенома, или это различие на комбинации взаимодействия геномно-эпигеномного уровня? Различия в заранее заданных настройках колок* через более точную регуляцию меток и инсертионем*? Условно можно говорить, что наши гены — это «железо» (Hardware), а эпигенетика — это «софт» (Software). Софт может быть коммитированным (детерминация через «окно возможностей») или привнесённым извне, например механизмами сенилита, да и вообще из вне многочисленными факторами. Это означает, что у мыши и землекопа «железо» может быть общее, а «софт» – разный. Софт – это механизм экспрессии и репрессии генов (включение – выключение, доступно - недоступно). Софт = эпигенетика — это, по сути, закладки, заметки на полях и маркеры выделения. Они не переписывают текст, но говорят клетке, что читать, а что игнорировать. Причём софт определяет целые большие комбинации (композиции) генов, определяющих ту или иную глобальную функцию клетки, задаёт путь, открывает «шлагбаум на развилках», направление жизнедеятельности клеток (гликолиз, сенесцент, апоптоз, реювентализация…). Но софт – это одновременно и КЭШ.
В чём различия кэша у разных видов и фенотипов? Очевидно у мышей софт сложился эволюционно под воздействием агрессивной среды, которая постоянно содержит организм мышей в стрессе и задача в первую очередь для вида – быстрая репродукция популяции. У землекопов условия среды иные: они живут в темноте при достаточном уровне качественных пищевых субстратов и полном отсутствии факторов стресса. Похоже, софт у землекопов представлен своим сводом коммитаций на эпигеноме (кодекс, правил игры многочисленных генов как единый ансамбль). И этот кодекс в нормальных условиях имеет большую значимость чем внешние факторы (сон, пища, стрессы, физическая активность…).
Как работают коммитации на эпигеноме? У разного типа клеток активны разные группы генов. Это и есть результат работы эпигенетики: направлять активацию тех групп (блоков) генов, которые определяют выбор пути нужного в конкретной ситуации. Таким образом, эпигеном это «управляющий штаб», координатор, направляющий на те или иные функции.
Такие «переключатели» запускаются с помощью:
- метилирования ДНК (маленьких химических меток, которые выключают гены),
- гистонов (белков, обёртывающих ДНК и управляющих доступом к информации),
- ферментов, реагирующих на сигналы из окружающей среды.
Интерес представляет
Определение причин различия продолжительности жизни у близких видов. Очевидно ответ на этот вопрос поможет понять эпигенетические механизмы продолжительности жизни. Рассмотрим частный пример с пчелами и термитами, у которых имеются различные фенотипы и касты в пределах одного вида. Казалось бы, один вид, один генотип - один фенотип, но у термитов и муравьёв 5 фенотипов. Это фенотипическое разнообразие, см. рис. 1.
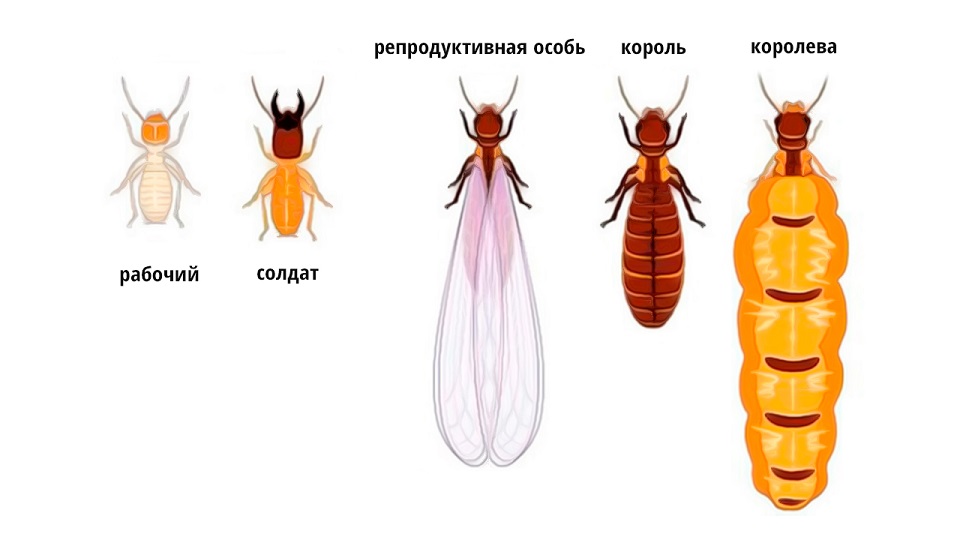
Рис. 1. На рисунке: фенотипические разнообразие особей среди термитов, которое существенно влияет на длительность онтогенеза.
У человека образование фенотипов зависит не от внешних факторов, а от этапов онтогенеза, возраста, внутренних факторов. Так например, имеется фетальные детские и юношеские фенотипы, которые существенно отличаются от взрослых с их бородами, гирсутизмом и маскулинностью. Это уже фенотип воина, рабочего. Не говоря уже о фенотипах старости… Такое же разнообразие происходит и при образовании модификатов иммунных клеток: М1, М2… Модификаты не связаны с морфогенезом (инициируется изнутри на уровне генов), а с фенотипическим разнообразием (инициируется на уровне эпигенома). Морфогенез – это последовательный процесс трансформинга, саморазворачивания системы. Фенотипия – это процесс координации на внешнем уровне со стороны эпигенома. Каким образом внешняя среда (факторы) запускает эти фенотипы? Пчелиные матки живут постоянно в темноте (очевидно важен этот фактор темноты), а главное они питаются особым пищевым субстратом – маточным молочком – источника вителлогенина. Так может быть это вещество является ключом для сбрасывания КЭШа? По сути, вителлогенин не просто активатор ювентальности, но и репликаций, а также полноценный пищевой субстрат. По сути, он играет роль фактора парабиогенеза – усилителя ювентальности, а значит и настройщика эпигенома на это? Похожими свойствами обладают препараты ДИЭНАЙ и им подобные на основе молок рыб. У других фенотипов пчёл имеется разнообразие активности и образа жизни, что тоже запускает комплекс генов иного фенотипа. Причём этот комплекс генов запускается с помощью эпигенома и его мантии, а они срабатывают на внешние эдафические факторы. У муравьёв из одного одновозрастного поколения может образоваться 5 фенотипов, которые внешне существенно отличаются
У растений настройки Кэша часто на много легче можно сбросывать. Так, например, агавы в природе живут около 15 лет. Они монокарпики, то есть один раз в жизни дают цветочную стрелку, плодоносят и затем гибнут, Но если у них удалять цветонос, то тем самым можно продлить жизнь до 80-100 лет. Следовательно их «железо» позволяет жить долго, а «софт» – этому мешает. Точно также сельдерей можно путём удаления цветоноса перевести из категории 1-летников, в категорию 2- и более летников. В данном случае причиной софта на укорочение жизни является выработка цветоносом и семенами гормона абсцизовая кислота, которая и влияет на особенности софта и эпигенома. Следовательно у человека тоже имеются регуляторы софта и ювентальности. Но они глубоко заглушены.
Таким образом, можно сделать предварительные выводы, что у человека мероприятия по сбросу КЭШа важно сочетать с методами аутофагии и полуавтономии на фоне привлечения факторов парабиогенеза.
Сенесцент и онкология – это дезинформация, груз, «мусор», которые появляются на уровне эпигенетики и ведущие к появлению новых неуправляемых автономно существующих модификатов, «паразитических» субъединиц. В тоже время известны факты хорошо задокументированных быстрых, практически «мгновенных» самопроизвольных излечений от самой тяжелой онкологии на 4-ой стадии – это яркие примеры сброса КЭШа – онкологической информации на эпигеномах клеток. Сбросьте КЭШ и не нужны будут ни какие операции, химиотерапия… Конечно, задачей становится в первую очередь очищать клетки именно от этих кешей. Эта дезинформация не только постоянно отбирает энергию у здоровых клеток и программ, но и постепенно глушит работу всего организма, подводя его к мортальному исходу.
Что понимается под сбросом КЭШа в клетках? В компьютерах сброс кэша осуществляется с помощью специальных программ, в т. ч. и антивирусных. В клетках кэш это вредная для них программа, статус. В природе многие виды легко сами способны сбросить ненужный им кэш и ювентализироваться. Примером этого могут быть каланхоэ, который способен образовывать на своих листочках ювенильные почки, а из них «детки» - новые молодые растеньица. Роща тополя осинолистного в возрасте 80 тысяч лет тоже способна легко возвращаться к ювенильности через подземные столоны (вегетативное саморазмножение). В животном мире таким примером может быть медуза, которая способна жить неограниченно, правда происходит это через преобразования в обратный метаморфоз в полип. Это означает, что она может сбросить груз зрелости и даже морфогенеза. В данном случае реморфогенез идентичен ювентальному реверсу. В итоге не особь, а один и тот же геном существует не лимитировано.
Чем отличается эффект сброса КЭШа от аутофагии? Автор понимает под понятием сброс КЭШа – возврат системы из состояния дисклаймера*, то есть патологического жизнеобеспечения за пределами гомеостаза, когда нарушены настройки на эпигеноме, с помощью механизма скадар* на МБК. Именно это и является целью метода полуавтономии на фоне парабиогенеза.
Принцип работы аутофагии основан на запуске механизма аутофагосом, с помощью которых происходит чистка клетки внутри цитозоля. Это естественный процесс «самоочищения» клеток, который реализуется с помощью специальных внутриклеточных структур, лизосомы, содержащие ферменты для расщепления «отработанного материала». При этом происходит слияние аутофагосомы с лизосомой. Их мембраны связываются, и ферменты лизосомы начинают расщеплять содержимое. Это позволяет клеткам организма избавляется от повреждённых или изношенных компонентов. Повреждённые белки и органеллы расщепляются и перерабатываются в энергию и строительный материал для новых структур, то есть рециклинг веществ.
Как происходит зарождение неопластических фенотипов (онкологических)?
Особенностью раковых клеток является то, что они находятся в гликолизе и при этом не могут воспользоваться кислородом даже в его присутствии. Это связано с глубокими вторичными подстройками в митохондриях к новым эдафическим условиям среды на фоне дисклаймера для хроматина (отсутствия уравновешенной взаимоподдержки между конформациями хроматина и корреляций с ним активности митохондрий). Каждый из них плохо сопряжен с деятельностью другой стороны, между ними информационный разрыв и каждый работает автономно. Одно не уравновешивает другое. Механизм маятникового балансирования и взаиморегулирования между ними сдвинут и не работает. Выход один: вернуть этот маятник в исходное состояние. Это можно осуществлять только реципрокными методами (мелкими шажками) подталкивания-выталкивания в норму той стороны рычага регулировок, который не отвечает (контрпозитный рычаг). Любые резкие движения с этим маятником заведут его еще в больший дисклаймер и уменьшат клиренс возможностей авторегулировок.
Этому способствуют затяжной клеточный автостресс и автопровоспаления. Клетки сами не могут выйти из этого состояния и сами себя провоцируют. Процесс разгорания гликолизности происходит по началу обычным путём, точно также как это происходит в норме у спортсменов при забегах и т.п. Здесь тоже естественным путём происходит деполяризация мембран клеток – недостаточно минусовых = «живых» зарядов. Это автоматически запускает процесс гликолиза. При этом митохондрии практически не работают, но при снятии нагрузки срабатывает система обратного сигналинга (контрпозитного) и вся система возвращается к норме и опять запускаются митохондрии. Регулировка происходит за счёт маятникового механизма сигналингов прямого (активирующего, транспозитного пути) и обратного (подавляющего, контрпозитного). Однако в случае затяжного запуска стрессового сигналинга транспозитов, система по причине глубоких провоспалений, не совместимых с требованиями гомеостаза, выходит за пределы нормируемых генетических констант (контроль уровня градиентов по ряду параметров), за пределы границ оптимума. Выход в «слепую» зону энтроукта, где система практически не регулируема и сигналинги сбиты. Это в первую очередь сказывается на работе МалогоБиоКристалла и его хроматина. Поскольку энергетика здесь в затяжном режиме находится в гликолизе и появляется огромное количество неопластических метаболитов со сменой секретома (SACanP — ассоциированный с канцером секреторный фенотип) происходят глубокие сбои настроек в хроматине. В этом случае по причине сбоя настроек на хроматине такая система становится нечувствительной (резистентной). Причем процесс дисклаймера (утраты ответственности за контроль и регулировку) происходит в хроматине в паттернах находящихся над блоком регулирующих генов (что делает их видимыми и экспрессируемыми), ответственных за весь фронт (блок) контрпозитных сигналингов. По этой причине митохондрии становятся неуправляемыми из-за недостаточности сигналинга со стороны хроматина. Процесс депривации сигналинга происходит не сразу, а постепенно и активность митохондрий постепенно тухнет путём ослабления кислородного метаболизма и усиления в помощь, компенсаторно гликолизного метаболизма. В этом случае гликолизная энергетика клеток осуществляется не в митохондриях, а снаружи их. При этом запредельный градиент SACanP (резко превышает требуемые константы) стимулирует мощный шквал провоспалений, а он является стимулом на пролиферацию. Неконтролируемые репликации происходят по причине того, что клетка воспринимает провоспаления как травму ткани и необходимость регенераций.
Затухание работы митохондрий ведёт к деполяризация мембран клеток. Митохондрии, кроме своих основных функций, еще поддерживают высокий потенциал зарядов как на ретикулярной сети клетки, так и в пределах ядра клетки. Этот же потенциал обеспечивает нормальные условия для хроматина, заданный его конформационный статус и конформацию его белков, а также мантии вокруг него, не допускает возможности сбоев в эпигенетических настройках.
Напомню школьный эксперимент с электростатическими лепестками, когда они заряжены оба минусовым зарядом, то в электроскопе происходит раздвижение пластинок, они зависают как в полёте. Точно также и хроматин в нормальном для него электропотенциале тоже способен расправляться, снимать напряжение и натяжку. Электропотенциал для него как смазка, в которой он находится как в левитационном состоянии, в невесомом плавании. В этом состоянии «левитирования» раскрываются его конформационные возможности в виде доступности хроматина для транскрипций. Это его нормальное состояние, которое и стремятся поддерживать механизмы гомеостазов, сверяясь и ориентируясь на константы, сравнивая реальные параметры с необходимыми с помощью прямых и обратных сигналингов, при необходимости их корректируя. Но в ряде случае система выходит за пределы этих констант и становится неуправляемой (невидимой, недосягаемой).
Такая система перестаёт выполнять свою миссию, не несёт свои обязанности (состояние дисклаймера). Это не поломка и не мутация – это дисклаймер!, когда не возможно выполнять свой морфогенетический алгоритм, у которого есть цель – выполнение миссии. Миссия становится невыполнимой. Как следствие – все многочисленные последствия в виде саморазрушений, но они не результат износа или старения – а последствия «забвения» обязанности, выполнения клаймера. Поддержка и сохранение клаймера – это самоцель системы, то есть её самосохранения. Система теряет цель (смысл) существования.
При этом особенностью сенесцента системы клеток в том, что они сами вводят себя в состояние дисклаймера, тогда как при онкологии это делают внешние факторы. Лечить такую систему бессмысленно, так как у неё нет цели к запуску на самовосстановление. Здесь нужен не ремонт от разрушительных последствий, а возврат цели = возврат к саморегулированию, вывод из цейтнота, когда сложную клеточную систему заклинивает, она впадает в забвение информационного плана, теряет «инстинкт» самосохранения. Цель поддерживать своё постоянство вокруг зоны констант и не допускать выхода за пределы, когда сложная клеточная система теряет координацию и общее направление развития, что приводит к деградациям в виде сенесцента или неоплазии – двум край-ностям одного процесса. Это два возможных варианта ответа на потерю ориента-ции, цели, когда сбит механизм ориентиров, наподобие тому как это происходит с прибором гироскопом у самолёта. Без гироскопа самолёт становится неуправляе-мым. Клетка без конформационного статуса хроматина тоже становится не управляемой, что = сбою в МБК. У самолёта есть заданный путь, цель, ориентир. В системе клеток организма тоже есть свои ориентиры навигации по Онтодианной Линии Жизни. Ориентир указывает как правильно вести себя клеткам в отношении их самообновления и пролифераций. В одном случае (перекосе крыла маятника) сбой может повести к утрате пролифераций (сенесцент), а в другом – к неуправ-ляемому хаотическому существованию, не поддерживающему их цель, смысл (неоплазия). В обоих случаях система становится неуправляемой, теряется её смысл, цель.
Ремодификация (восстановление в норму у онкоклеток) и реювентализация (у сенсцентных клеток) – означает не только устранение митохондриальных дисфункций и восстановление аэробной энергетики, но и сброса КЭШа на хроматине с последующим восстановлением конформации хроматина. Причём КЭШ может быть разным, с диаметрально противоположными свойствами: специфическим (локальным и присущим для неопластических процессов) и неспецифическим (тотальным, присущим для процессов сенесцента). Митохондриальные дисфункции и КЭШ на хроматине взаимозависимы и в запредельных случаях создают «патологические кольца», когда невозможно корректировать восстанавливать систему не разорвав эти кольца взаимозависимости.
Если клетка не способна ремодифицировать, даже при исчезновении причин, вызвавших проблему анаэробного гликолиза, то она должна уйти на апоптоз. Но у раковых и сенесцентных клеток механизм возврата с анаэробного в аэробный путь энергетики «заклинивает» не только из-за слишком глубоко зашедших перестроек-настроек в эпигеноме, но и кризиса конформаций эпигенома, а также изменений в мантии и невозвратных перестройках в митохондриях. Все вместе они становятся помехами не просто на пути возврата в норму, а даже к апоптозу. Гены апоптоза находятся в митохондриях и становятся недосягаемыми для участия в регулировках. Такие изменения уже носят не функциональный, а структурный характер (нерегулируемый, не регламентированный правилами). В таком случае для запуска апоптоза нужны более жесткие!!! условия и требования для их выбраковки, которые, очевидно, возможно осуществить в условиях полуавтономного режима, где ещё возможно запускать реципрокные шажки регулировок. Но будет ошибочным считать что полуавтономия – это и есть способ лечения. Нет, полуавтономия создаёт лишь условия для реализации других реципрокных методов и мягкого возврата к норме системы балансиров и буферов, а точнее к снятию конформационного дисбаланса на хроматине. Это запустит гироскоп самоуправления и самосохранения, то есть вернет клеткам их цель и смысл. При этом, предполагалось, что в первую очередь пострадают менее податливые и менее гибкие к приспособлениям клетки (резистентные). Основывалось это предположение на том, что высокая чувствительность и дифференциация клеток может быть только у аэробного типа клеток, тогда как онкологические и сенесцентные клетки не отзывчивы, теряют значительную часть дифференциации и анаэробны. Но эта позиция опровергается тем, что голод и гликолиз наоборот делает онкоклетки более агрессивными. Такой же негативный сценарий и для процесса сенесцента.
Итак, раковые клетки менее прихотливы и более агрессивны, их конкуренция за выживаемость выше чем у здоровых. Во время голодания или без него в приоритете для получения пищевых субстратов всегда будут раковые клетки. Даже если онкобольной человек голодает, то страдает всегда организм весь, а раковая опухоль как более агрессивная всегда будет перехватывать и отбирать пищевые субстраты и продолжать расти. Опухоль растёт, процветает и доводит организм человека до кахексии (полного физического истощения и краха). Это объясняет почему онкобольным нельзя голодать. Тогда возникает вопрос почему при онкологии автор допускает полуавтономию, ведь это тоже полуголод? Задача полуавтономии не заморить голодом онкоклетки, не довести процесс до крайности и усугубить его, а сбросить КЭШ, груз который позволяет убрать дисклаймер* (диссонанс в регулировках), оголить защиту раковых клеток, сделать их менее агрессивными.
Возможные механизмы положительного содействия полуавтономии в противодействии раковому состоянию. Очевидно содействие этому будет осуществляться через форсирование процессов аутофагии, уменьшение градиента токсического выброса секретома SACanP*, который является условием для самостимулирования раковых клеток. Автор считает, что это в сочетании с другими методами позволит запустить скадар* в МаломБиоКристалле, поможет убрать митохондриальные дисфункции, а затем откроет возможность восстановить конформацию хроматина и восстановит хроматиновые настройки. Для этого в организме нужно создать особую эдафическую среду, когда «больные» онкофенотипические клетки смогут запустить механизмы репарации хроматина. Отмечу, что в условиях обычного организма такую среду создать невозможно, так как требования к репарации повреждённого хроматина у онкоклеток далеко не совместимы с требованиями самого организма пациента. Поэтому здесь нужны методы эквилибристики, компромиссов, чтобы выталкивать онкоклетки из своего статуса -онкофенотипа и возврата в статус нормальных клеток. В эти требования входит:
- восстановить аэробную работу митохондрий,
- ослабить секретом SACanP,
- устранить деполяризацию на мембранах клетки,
- запустить скадар в МаломБиоКристалле
- изменить повреждённую конформацию хроматина со смещением зон видимости и невидимости генома,
Весь перечисленный комплекс возможных мер для ремодификации онкоклеток и реювентализации сенесцентных очень похож на подведение этих клеток к состоянию близкому у здоровых клеток, вывести их из глубокого цейтнота, а точнее зоны энтроукта* (далеко за пределами возможности регулировок гомеостазов), то есть неуправляемости, что и позволит запустить механизмы репараций или апоптоза. Подобные благоприятные условия, очевидно, существуют только на стадии эмбриогенеза, где заряды мембран, например, достигают -200 мв, тогда как у обычных клеток -70 мв. Именно на этапе эмбриогенеза доминируют протопрограммы клеток (древние, базовые), но заторможены примапрограммы (более поздние надстройки, отвечающие за дифференцировку).
Диаметрально противоположная направленность дисклаймеров* у раковых и сенесцентных клеток. Известно, что в раковых клетках наблюдается дисбаланс метилирования ДНК, который проявляется в двух ключевых типах изменений: глобальном гипометилировании и локальном гиперметилировании промоторов генов-супрессоров опухолевого роста. Это сопряжено с разной натяжимостью хроматина по всей его длине, а значит одни зоны перенатянуты, а другие наоборот - чрезмерно компактизированы. Это и есть сбой настроек хроматина. Причём у сенесцентных и раковых клеток этот дисбаланс имеет диаметрально противоположную направленность. Это означает, что у сенесцентных клеток в отличие от раковых будет происходить глобальное гиперметилирование и локальное гипометилирование, что в любом случает нарушит настройки взаимодействия между прото- и примапрограммами. В любом случае будут проявляться крайние формы разрегулировок: от полной остановки репликаций до не лимитированных неуправляемых репликаций.
Условия необходимые для возврата сенесцентных и раковых клеток в зону клаймера* (оптимума, гармонии). В таких случаях надо создать условия, когда клетки смогут автоматически вернуться в зону клаймера* (ответственности, гарантии регулировок), восстановить натяжку хроматина, его конформацию и привести в норму его метилирование. Это означает запустить работу колок – инсертионем* (меток) и их инсертионемаз (ферментов регулирующих колку). Важно отметить, что теломеры у раковых клеток удлинены, а значит гипертрофирована работа теломеразы, над ней нет регулирующего контроля, она работает вразнос. Это полностью противоположно тому, что происходит у сенесцентных клеток, у которых теломеры наоборот укорочены (блокирована теломераза). Гипертрофирована – блокирована работа теломеразы, а ведь это яркий пример разных типов сбоев работы настроек на хроматине. Регулируют не теломеры хроматин, а наоборот хроматин теломеры. Механизм укорочения или удлинения теломер иной чем считалось и это следствие конформационных перестроек хроматина, а не причина. Точно также идёт сбой регулировок и по всей остальной длине хроматина в его инсертионемах. Хроматин «дубеет», становится нечувствительным к регулировкам, типичная хроматиновая резистентность. Это еще раз говорит о тотальной разбалансировке настройки хроматина при раке и сенесценте, но с противоположной направленностью. Сенесцент и рак – это два крайних состояния, два крыла одной маятниковой системы регулировок на хроматине. Они определяют выход автосистемы регулировок за пределы оптимума, а точнее в область неуправляемости. Между ними находится точка оси, а точнее точка константы, определяющая параметры нормы. Вокруг этой точки находится коридор зоны нормы. Он определяет границы оптимума где возможно поддерживать гомеостаз, за этими границами начинается область хаоса, энтроукта.
Восстановление рН и буферной среды онкоклеток с помощью минералов Mg и др.
Задача не просто устранить SACanP и восстановить рН и буферную среду вокруг онкоклеток, а значительно превысить нагрузку на противоположной рычаге маятника, что бы «раскачать» систему. Это означает что автор предлагает поднимать рН в строну ощелачивания на много выше чем требуется в норме. Особым благоприятное содействие видит в применении минералов в критически максимально высоких дозах, в частности минерала Mg в цитратной форме в виде препарата «Магний Экспресс Форте» от ВИТАУКТ.

А еще лучше под названием «Ренимарен» или «Магний Тройчатка».
или

Высокая минерализация позволит легче и дольше переносить полуавтономный голод. При голодании в первую очередь критическим становится нехватка комплекса минералов, а не органических веществ. Для клетки важны 4 «столбовых» макроминерала, на которых держится гомеостаз, это: Mg, Na, K, Ca. Но особо выделяется значимость Mg. На минералах организм может в десять раз дольше переносить период голодания. Нехватка минералов является большим стрессом, чем органики. О значимости Mg при онкологии смотрите книгу и статью автора [2, 3]. Магний один из соучастников процессов по снижению уровня провоспалений и коррекции рН, что особенно важно при онкологии. Локальное увеличение концентрации магния в опухолевой ткани:
- усиливает ответ Т-клеток,
- будирует работу митохондрий,
- противодействует гликолизу,
- влияет на уровень витамина D,
- подавляет канцерогенез,
- подавляет развитие опухоли за счет ингибирования глутатиона,
- угнетает рост карциномных опухолей,
Но сам по себе Mg не в состоянии отрегулировать онкологический статус и привести к апоптозу или ремодификациям. Здесь нужен целый комплекс мер, включая противовоспалительный D3, мелатонин, запуск аутофагии…
Аутофагия в роли как супрессора опухоли, так и повышения устойчивости к другим методам лечения. Аутофагия может как содействовать, так и противодействовать методам лечения опухоли. Почему? Если мы воздействуем на опухоль мягко, то ограничиваем её потенциал и ведём к перенастройкам, ремодификациям, выводим из зоны цейтнота, если действуем жёстко – то повышаем агрессивность опухоли, усиливаем механизмы борьбы за выживание. При этом степень гликолизности онкоклеток повышается, а аэробная активность митохондрий понижается.
Двойственность возможных путей ответов онкоклеток при аутофагии связа-на с её способностью влиять на клеточный гомеостаз, выживание клеток и меха-низмы резистентности к терапии. Это зависит от типа клеток, стадии опухолевого процесса, условий окружающей среды и других факторов.
Механизмы двойственной роли аутофагии:
1. Содействие выживанию опухолевых клеток и их адаптации к стрессу. Аутофагия позволяет раковым клеткам выживать в условиях дефицита питательных веществ, гипоксии, окислительного стресса или под воздействием противоопухолевых препаратов. Она обеспечивает переработку внутриклеточных компонентов, что поддерживает энергетический баланс и процессы биосинтеза, необходимые для роста и выживания клеток. Например, в гипоксических зонах опухоли аутофагия помогает клеткам адаптироваться к неблагоприятным условиям.
2. Подавление опухолевого роста на ранних стадиях. В некоторых случаях аутофагия может действовать как механизм, подавляющий опухолевый рост, предотвращая накопление повреждённых органелл и белков, что снижает вероятность малигнизации клеток и замедляет развитие опухоли. Например, на ранних стадиях развития опухоли активация аутофагии может приводить к гибели раковых клеток.
3. Влияние на метастазирование. С одной стороны, аутофагия может обеспечивать энергию и питательные вещества для эпителиально-мезенхимального перехода (ЭМП), который способствует распространению метастазов. С другой стороны, на более поздних стадиях аутофагия может подавлять колонизацию метастазов.
4. Связь с резистентностью к лечению. Аутофагия может способствовать формированию лекарственной устойчивости, поддерживая выживание раковых клеток в состоянии покоя или старения в ответ на терапию. Также она может влиять на переход раковых клеток в состояние покоя, которое выживает после химио- или радиотерапии.
Влияние на агрессивность опухоли и метаболизм:
- Повышение гликолизности онкоклеток может быть связано с адаптацией к стрессу: в условиях гипоксии или недостатка ресурсов клетки могут переключаться на анаэробный метаболизм (гликолиз), что повышает гликолитическую активность. Аутофагия может поддерживать этот процесс, обеспечивая необходимые метаболические субстраты.
- Понижение аэробной активности митохондрий также связано с ролью аутофагии. Митохондрии играют ключевую роль в аэробном дыхании, и их дисфункция может снижать аэробную активность. Аутофагия может влиять на состояние митохондрий, например, через селективное разрушение митохондрий, что снижает образование побочных продуктов, повреждающих ДНК и способствующих мутациям.
Терапевтические стратегии:
- Ингибирование аутофагии может повышать чувствительность опухолевых клеток к противоопухолевым препаратам, преодолевая лекарственную устойчивость. Например, использование ингибиторов аутофагии в комбинации с химиотерапией может усиливать цитотоксический эффект.
- Активация аутофагии в некоторых случаях может повышать чувствительность раковых клеток к химиотерапии. Однако важно учитывать контекст и стадию заболевания.
Таким образом, роль аутофагии в онкологии не однозначна и требует индивидуального подхода к терапии с учётом конкретных молекулярных механизмов, типа опухоли и состояния пациента. Дальнейшие исследования в этой области направлены на разработку более точных методов модуляции аутофагии в рамках терапевтических стратегий. Из за этой неоднозначности становится невозможным доводить лечение до полного конца. Метод позволяет лишь в определённой степени сдерживать процесс, но дальнейшее его форсирование приводит к обратной реакции.
Как убрать эту вторую фазу обратной реакции опухолевых клеток? Известно, что обратный процесс противодействия усиливается при повышении доз воздействия. В этом случае опухоль становится или более агрессивной, или уходит на некроз, что пагубно сказывается на всём организме. Это означает, что лечебный процесс сдерживающего действия на опухоль возможен только в ограниченной зоне воздействия, а логика говорит, что в этом коридоре возможностей и надо воздействовать, постепенно расширяя коридор, отвоёвывая всё новые пространства. Но в чистом виде этот приём позволяет улучшить качество жизни онкобольного, ослабить онкосимптоматику и продлить ему жизнь, но не позволяет полностью устранить опухоль, она просто будет развиваться медленнее.
Мягкие реципрокные методы комплексных мер по полной ремодификации раковых клеток. Теоретически это самый идеальный подход в лечении рака. Автор считает наиболее правильным и гуманным разрабатывать это направление лечения. При этом комбинация аналогичныхметодик имеющих синергию, однонаправленное действие расширяет диапазон влияния, коридор возможностей и усиливает их лечебный потенциал.
Поэтому автор утверждает, что эффект от метода «полуавтономии» можно значительно усиливать путём сочетания с синергичными методами. Цель – вывести мягко и без побочки раковые клетки в зону клаймера и длительно удерживать их там. Это позволит подключиться механизмам автонастроек на эпигеноме и авторегулировок в клетках. Другими словами клетки восстановят свою цель, смысл.
Особенностью метаболизма онкологических и сенесцентных клеток является ложный гликолиз и это их уязвимое место. Как раковые, так и сенесцентные клетки существуют в основном за счет гликолиза, причём отличия их лишь в разной глубине гликолиза. Не является ли это явной подсказкой направления воздействия на них для трансформации в нормальные клетки?
В норме переход в гликолиз регулируется параметрами уровня кислорода и отношения уровня АТФ к АДФ и АМФ, которые служат аллостерическим регуля-тором скорости гликолиза. Накопление АДФ и АМФ (при потреблении АТФ) стиму-лирует гликолиз, а высокий уровень АТФ — подавляет.
Полуавтономия создаёт лишь благоприятный фон, условия для подключения комплекса других методик запускающих апоптоз или ремодификацию. Дальше уже создаются благоприятные условия для других методик, в том числе уже слабой безвредной химиотерапии, методам парабиоза*, а также для активирования аэробной энергетики митохондрий и восстановления значимости эффективности антиоксидантов и антиинфламантов (противовоспалительный эффект).
Антиоксиданты и антиинфламанты при онкологии могут играть двойственную роль: в одних случаях они сдерживают онкопроцесс, а в другом – наоборот его провоцировать. Всё зависит от их дозы. Следует учитывать, что раковые клетки находятся за пределами гомеостазов и то, что для здоровых клеток может оказывать положительное действие, для раковых будет работать в обратном направлении и всего лишь разжигать улучшать работу гликолиза. Маленькие дозы не работают, а большие – провоцируют рак. Напомню, что сбросить КЭШ – это означает успокоить, ослабить провоспаление и гликолиз. На этом фоне правильно заработают антиоксиданты и антиинфламанты.
По сути, антиоксиданты - это «полиция» или «пожарная команда». Они нейтрализуют свободные радикалы, отдавая им свой электрон. Но это правило работает только при низких дозах, а при повышении доз эффект переходит в противоположный – оксидативный стресс, который в онкоклетках итак имеется в повышенном уровне, а дополнительное усиление стресса приведёт к усилению в ней ангигенеза (рост сосудов) и повышению агрессивности опухоли, в крайнем случае к её некрозу. Антиоксиданты активируют белок BACH1, который заставляет опухоль переходить на аэробный гликолиз (эффект Варбурга) - именно это стимулирует метастазирование.
Особенно велик уровень оксидативного стресса у солитарных крупных опухолях, дополнительный гиперстресс от гипердоз антиоксидантов может ухудшить статус локализованной опухоли, но при этом усилить её склонность к метастазированию, мигрированию раковых клеток. Миграция клеток – это закон природы: когда среда для существования становится не оптимальной, начинают срабатывать эволюционно древние принципы закрытых систем, ухудшающих статус колонии и провоцирующих её на миграцию, если миграция не удаётся, то такая система постепенно отмирает. Но отмирание идёт по механизму некроза и интоксикаций, что приносит порой еще больший вред организму.
Точно также как антиоксиданты действуют и активаторы энергетики клеток или по другому - так называемые прекурсоры NAD+. Самая известная из них - никотинамид рибозид (NR), форма витамина В3. Её рекламируют как средство от старения, для энергии и восстановления ДНК. Изучение влияния никотинамид рибозид на раковые клетки дало плачевные результаты: оказалось, что NR способен стимулировать рост и, что ещё страшнее, метастазирование рака. В частности, повышается риск развития «трижды негативного рака молочной железы» - самой агрессивной формы, у которой нет трёх основных рецепторов (эстрогена, прогестерона и HER2), что делает её почти неуязвимой для стандартной гормональной терапии. Кроме того, витамин В3 в этой форме стимулировал образование метастазов в головном мозге. Почему? Никотинамид рибозид повышает уровень молекулы NAD+ в клетках. NAD+ - это топливо для множества процессов, включая энергетический обмен. Раковая клетка - ненасытный потребитель энергии. Дополнительное топливо она использует не для того, чтобы стать «здоровее», а чтобы быстрее делиться и мигрировать по организму.
Витамин В12 в роли провокатора рака. Конечно в этом случае обсуждаются избыточные его дозы. Витамин B12 поддерживает рост клеток в целом — не только здоровых, но и потенциально предраковых. Обнаружена U-образная зависимость между потреб-лением B12 и риском рака: повышенный риск наблюдался как при дефиците вита-мина, так и при его избытке. Но это показывает лишь статистическую связь, а не прямую причинно-следственную зависимость. То есть действие может быть кос-венное, опосредовано через другие механизмы.
Обнаружено, что производные витамина В3 (NMN, NR, NAM) снижают эффективность химиотерапии при раке поджелудочной железы.
Автор ожидает, что полуавтономия создаст благоприятные условия для проявления эффективности таких препаратов как Mg-цитрат, янтарная кислота, NMN (Никотинамидмононуклеотид), Фуллерен С60, D3, Куркумин-Адванс, мелатонин и др. в невысоких дозах (безопасных, защитных).
Почему официальные методы лечения рака во многих случаях заканчиваются метастазированием или рецидивом? Более 75% рецидивов случаются в первые пять лет после терапии. Научный консенсус долгое время сводился к простой идее: «недобитые» раковые клетки снова начинают расти. Но так ли это? Оказалось, что само лечение может невольно создавать условия для нового витка болезни, создавать почву для метастазов, вызывать в организме изменения, благоприятные для распространения рака. Принципы и цели химиотерапии - убить раковые клетки, что по сути аналогично методу гипероксидативного стресса, как способа лечения для онкоклеток. Но это коварный варварский метод, так как любой оксидативный стресс – это путь к сенесценту. Даже такие безобидные и полезные вещества как антиоксиданты, витамины, антиинфламанты при завышении доз тоже ведут к тому же оксидативному стрессу и могут провоцировать рак и сенесцент. Тот же оксидативный стресс якобы как лечит, так и провоцирует эту же болезнь. Точно также очевидно действуют препараты от старения и сенесцента: до поры до времени они в определённой степени сдерживают процессы, а потом (на высокой степени его развития) начинают сами же его провоцировать, содействовать!
Высокие дозы как химиотерапии, так и оксидантов в большинстве случаев действует как шок как для больных клеток, так и здоровых, различия лишь в разной степени этого шока. Шок + КЭШ = скрытая память болезни. Причём чем выше стадия развития болезни, тем выше вероятность её рецидива после лечения.
Автор считает, что причиной рецидивов как рака, так и сенесцента является остающийся КЭШ на хроматине клеток. Поэтому правильнее говорить о необходимости не просто уничтожения больных клеток (раковых или сенесцентных), а сброса полностью всего КЭШа, эпигенетической патологической памяти!
Эти препараты, а также рукопись книги «ЭПИГЕНЕТИКА МОЛОДОСТИ», можно заказать на сайте Гарбузова.
Термины
— Делеция — разрыв на хромосоме из-за нарушения натяжки и компактизации её в связи с проблемами на хроматине.
— Дисклаймер — это критические условия работы самонастраивающейся эпигенетической системы за пределами гомеостаза, где система регулировок статусов хроматина не имеет возможности нести ответственность за обеспечение гарантии стабильности, то есть работает не по правилам, в диссонанс. Это зона хаоса и свободы для энтроукта (разрушительные процессы), которые вынужденно, ошибочно запускают зоны хроматина ответственные за настройку работу генов обеспечивающих активность высокопотентных стволовых клеток (ювентальность). Также благодаря дисклаймеру сбой регулировок в противоположных частях эпигенома приводит к сенесценту. Очевидно эволюция направленно избрала механизм дисклаймера для запуска сенесцента. — Термин предложил Г. А. Гарбузов
— Клаймер — это зона оптимума, то есть благоприятных условий для работы самонастраивающейся эпигенетической системы в пределах гомеостаза, где система регулировок статусов хроматина имеет возможности обеспечивать гарантии стабильности, то есть работает по правилам, в резонанс.
— Колки — дословно это ключ-механизм настраивающий натяжку струны, то есть отвечающий за настройки инструмента, например гитары. В хроматине под колками подразумеваются механизмы поддерживающие степень натяжки хроматина. Таким образом, эти механизмы, по сути, являются системой настройки хроматина. — Термин предложил Г. А. Гарбузов.
— Инсертионемы — дословно означает вставка и подразумевает вставки в хромосомах, которые регулируют её натяжку и тем самым обеспечивают видимость или невидимость стволовых клеток. Действуют и регулируются подобно теломерам, которые регулируются активностью фермента теломераза. От степени метилирования и натяжки хромосом теломераза может быть заблокирована. У инсертионем имеются свои аналоги «теломеразы» - инсертионемазы. – Термин предложил Гарбузов Г. А.
— Парабиоз, в т. ч. информационный парабиогенез — от слов «биогенез», которое в эмпирическом обобщении означает стимулирование живого организма, а точнее когда одно живое активирует, оживляет другое, где «пара» (дословно — рядом, около) означает, что активация происходит за счёт другого живого начала. Информационный парабиогенез - воздействие информационной модели в виде её голографии, но не непосредственно на реальный биологический объект, а на Голограмму его Малого Биокристалла. — Термин предложил Г.А. Гарбузов.Такое понимание вопроса подразумевает, что считанную информацию в виде голограммы можно скопировать на носители и переносить, проецировать её на другой объект, например в ночное время, что вызовет эффект информационной когеренции в Малом Биокристалле. Следует отличать от термина Парабиоз или гетерохронический парабиоз — когда сшивают реальные кровеносные системы молодой особи мыши со старой, при этом происходит омоложение старой и увеличение продолжительности жизни, но состаривание молодой.
— Скадар — устройство для регулировок в состоянии эпигенома с помощью голограммных развёрток. Предназначен для регулировок процессов происходящих во времени и продвижения по Онтодианной Линии Жизни. Состоит из слов: «ска» от слова сканер и «дар» от слова радар. — Термин предложил Г. А. Гарбузов.
— Энтроукт — от слов энтропия и сила. Ослабление возможностей Витаукт, когда гармония всех систем организма выходит за пределы коридора оптимума нужного для гормезиса. - Термин предложил Г.А. Гарбузов.
— SACanP — ассоциированный с канцером секреторный фенотип. — Термин предложил Г. А. Гарбузов.
Книги автора
ЭПИФИЗ — ВЕРХОВНЫЙ ДРАЙВЕР ОНТОГЕНЕЗА. КЛЮЧ К МОЛОДОСТИ И СТАРЕНИЮ

Старение — это многоуровневый процесс как на верхних этажах регулировок (системном), так и на нижнем клеточном. Чтобы адресно разрабатывать методы противодействия и сдерживания общего потока старения (геронтос), необходимо найти правильный ответ — откуда исходят именно истинные первичные начала этого общего процесса. Автор провел глубокий анализ природы старения как в растительном мире, так и в животном. Показано, что этот процесс является двусторонним: снизу он исходит с уровня клеток (сенесцент), причиной является эпигенетическая коммитация на основе онтофизиса; а сверху верховным драйвером этого процесса является эпифиз, а не гипоталамус, как это считалось ранее. Исходя из этих фундаментальных разработок, предложен комплекс методик воздействия на все уровни сдерживания старения. Показана динамика взаимовлияния друг на друга верхнего и нижнего этажей старения. Именно верхний этаж определяет весь спектр возрастзависимых заболеваний (анизомалий). Если мы научимся сохранять «эпифизарную молодость» = неотению, то отодвинем старость.
ВЗЛОМ ИММУННЫХ МЕХАНИЗМОВ СТАРЕНИЯ — ПУТЬ К ЮНОСТИ И ЗДОРОВЬЮ

Эта книга знакомит читателя с принципами и методиками противодействия старению. Автор впервые разработал интегральную теорию старения организма, которая рассматривает его в виде взаимодействия двухуровневого процесса как на уровне клеток (сенесцент), так и на уровне всего организма (сенилит). Ознакомившись с материалами, приведёнными в данной книге, вы поймёте, что надо делать, чтобы как можно дольше сохранять молодость, здоровье и бодрость духа.
ЗДОРОВЬЕ, МОЛОДОСТЬ, ДОЛГОЛЕТИЕ. КАК МАГНИЙ И ОКСИД АЗОТА ПРОТИВОСТОЯТ СТАРЕНИЮ

Глубокий анализ существующих причин старения позволил автору создать свою единую теорию «биологии старения», которая строится на интеграции процессов сенесцента (на уровне клеток) и сенилита (на системном уровне). Показана решающая роль магния и оксида азота в регулировке на клеточном уровне сенесцента. Именно сенесцент (нарушение гомеостазов) определяет механизм сенилита (нарушение гормезиса и витаукта), а с ними и возрастзависимых «болезней», которые на самом деле не болезни, а естественные анизомалии, то есть механизмы реализации старения. Только правильное понимание сути природы старения позволит выйти на правильные пути преодоления возрастных болезней и продления жизни человека через механизмы неотении (продлённой молодости), чему посвящена следующая книга автора «Биология молодости».
САМОУНИЧТОЖЕНИЕ РАКОВЫХ ОПУХОЛЕЙ. ФЕНОМЕН АПОПТОЗА

Книги можно заказать на сайте garbuzov.org.
Онкология. Можно ли её вылечить безвредными способами? Автор впервые показывает, что можно и нужно! Для этого нужно запустить механизмы избирательного самоуничтожения раковых клеток в организме на основе клеточных программ апоптоза и аутофагии, которые нужно адресно открыть. Предложена Комплексная Программа из противоположно направленных методов, вычлененных в две фазы и направленных на расшатывание и перебалансировку основных жизнеобеспечивающих механизмов клеток, в т. ч. антиоксидантной « прооксидантной, провоспалительной « антивоспалительной и др. У онкоклеток коридор возможностей самозащиты, их гормезис намного уже, чем у нормальных клеток, у которых Жизненная Сила, Витаукт, намного выше. Такая разность возможностей позволяет создавать особые условия, когда становится возможным запуск механизмов самовыбраковки онкоклеток.
БИОЛОГИЯ МОЛОДОСТИ – электронная рукопись 2026 г

Ознакомиться с ней можно на сайте garbuzov.org.
Практически все авторы научно-популярных работ по анти-эйдж теме рассматривают всевозможные методы сдерживания старения и продления жизни, здоровья… Автор книги впервые показывает, что это не эффективно, так как не убирает суть проблемы и связывает эту тему через продление молодости, которая и есть фундамент для всего остального. Без продления молодости невозможно радикального решения всех остальных проблем. Автор подробно анализирует биологию молодости, объясняет почему она уходит. Важнейшими для запуска старения являются механизмы неспецифического иммунитета, а также старение клеток (сенесцент). Изучен весь мировой опыт и знания по продлению молодости на всех уровнях организма, что автоматически отодвинет все «болезни» старости - анизомалии.
Книгу или Программу-консультацию по заболеваниям Вы можете приобрести по нашему адресу.
Вы можете обратиться с вопросами или за консультацией к Гарбузову Геннадию Алексеевичу на сайте garbuzov.org, по адресу: vitauct@yandex.ru или по телефону 8 (928) 239-13-64.
Список литературы